서 론
전자담배는 2003년 개발된 이 후, 전세계적으로 널리 유통되고 있다. 많은 흡연자들이 일반담배(tobacco cigarette)에 비해 안전하다 하여 금연을 목적으로 많이 사용되고 있다. 하지만 안전성과 금연에 대한 효과에 대한 논란이 커지고 있으며 전자담배 사용에 따른 인체의 유해성에 대한 관심이 높아지고 있다[1]. 전자 담배는 태워 피는 일반담배에 비해 건강에 해롭지 않다고 알려져 많은 사람들이 일반담배 대용으로 많이 사용하고 있으나 니코틴을 포함하여 사용하는 경우 니코틴부산물에 대한 위험성은 여전히 남아있다[2]. 니코틴을 포함하여 사용하는 전자담배에서 다양한 담배특이니트로사민(tobacco-specific nitrosamines, TSNAs)이 발생하는데 대표적으로 5종류가 있다; N’-Nitrosonornicotine(NNN), 4-(N-Nitrosomethylamino)-1-(3-pyridyl)-1-butanone(NNK), N’-Nitrosoanabasine(NAB), N’-Nitrosoanatabine(NAT), 4-(N-Nitrosomethylamino)-1-(3-pyridyl)-1-butanol(NNAL)[3,4]. 전자담배나 일반담배 흡연이 인체에 장기간 노출될 경우 TSNAs과 같은 물질들에 의해 혈관 질환, 천식, 만성폐쇄성폐질환, 당뇨병 및 태아발달 등의 심각한 문제를 야기할 수 있다[5]. 또한, NNN과 NNK는 Group I(Carcinogenic to humans)으로써 실험동물에서 폐암, 식도암, 췌장암 등 여러 악성 종양을 유발한다고 알려진 강력한 발암물질이다[6]. 하지만 염증성 호흡기 질환에서 점액 과분비에 미치는 TSNAs의 영향에 대한 연구는 거의 없다.
점액은 기도 내부의 상피세포층을 덮고 있는데, 이는 외부에서 유입되는 유해인자들을 포집하고 점액섬모 청소기능(mucociliary clearance)을 통해 체외로 제거하여 기도 내부를 보호하고 기도 내 항상성을 유지하는 역할을 한다[7,8]. 점액은 표면에 위치하는 겔(gel)층과 섬모주위액층의 졸(sol)층으로 존재하며, 구성 성분으로는 97%의 수분과 1~3%의 지방, 탄수화물, 핵산, 당, 당단백질, 아미노산 등으로 구성되어있다. 그 중, 점액유전자는 분자량이 매우 큰 당단백질로서 점액의 점성과 탄성을 결정하는 중요한 성분이다[9]. 점액유전자는 크게 두 가지 형태로 존재하는데, 분비형 점액유전자와 막성 점액유전자로 구성된다. 대표적으로 호흡기 질환과 관련된 분비형 점액유전자로는 MUC5AC와 MUC5B가 있다[10,11]. 이러한 점액유전자는 활성산소와 다양한 사이토카인(Interleukin 1β, 6, 8 등)에 의해서 조절되며[12-14], 디젤배출입자(diesel exhaust particle), 카드뮴(cadmium)과 미세먼지(particulate matter)같은 환경오염물질에 의해서도 조절된다[15,16]. 또한 담배관련 주된 부산물인 acrolein은 호흡기상피세포에서 gelatinases와 관련 있는 MMP9을 증가시켜 MUC5AC를 증가시킨다고 보고되었다[17].
담배특이니트로사민은 발암물질로 암과 연관된 연구결과는 많이 알려져 있지만 염증성 호흡기 질환의 주요 병인 중 하나인 점액 유전자 과분비에 대한 연구결과는 알려지지 않았다. 따라서 본 연구는 담배특이니트로사민이 호흡기상피세포에서 점액유전자 발현에 미치는 영향에 대해 확인하였고 그와 관련된 수용체를 알아보고자 하였다.
재료 및 방법
재 료
N’-Nitrosonornicotine(NNN), 4-(N-Nitrosomethylamino)-1-(3-pyridyl)-1-butanone(NNK), N’-Nitrosoanabasine(NAB), N’-Nitrosoanatabine(NAT), 4-(N-Nitrosomethylamino)-1-(3-pyridyl)-1-butanol(NNAL)은 Toronto Research Chemicals(Toronto, CanadaSigma(St. Louis, MO, USA)에서 구입하였다. RPMI 1640 medium은 Invitrogen(Carlsbad, CA, USA), fetal bovine serum(FBS)은 Hyclone Laboratories(Logan, UT, USA)에서 구입하였다. MUC5AC(ab3649)와 MUC5B(ab77995) 일차항체는 Abcam(Cambridge, England, United Kingdom)에서 구입했고, HRP-conjugated 이차항체는 NOVUS Biologicals(CO, USA)에서 구입하였다.
세포 배양 및 처치
사람 폐의 점액상피양 암 세포주(human pulmonary mucoepidermoid carcinoma cell line)인 NCI-H292 세포(American Type Culture Collection, Manassas, VA, USA)를 6-well plate에 1×106 cells/well를 2 mM L-Glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin과 10% FBS가 포함된 RPMI 1640 배지를 이용하여 95%의 산소와 5%의 이산화탄소가 혼합된 배양기에서 37°C의 온도로 배양하였다. NAB, NAT, NNN, NNAL과 NNK이 기도 점액에 미치는 영향을 알아보기 위해서 NCI-H292 세포에 농도별 처리하고 점액유전자 발현과 단백질 생성을 유도시켰다. 대조군은 동일한 시간 동안 배지에서 단독으로 배양하였다. NCI-H292 세포의 세포독성 유무는 WST-1 분석(EX-Cytox, Innotech, Korea)과 현미경을 이용한 세포 형태 변화 유무를 확인하여 검증하였다. 이 연구는 본원임상 시험 심사 위원회(Institutional Review Board)의 승인을 받아 시행하였다(YUMC2019-08-057).
Real-time PCR 분석
Gene Amp RNA PCR core kit와 iQ SYBR Green Supermix(Bio-Rad, Hercules, CA, USA)을 T100TM Thermal Cycler와 CFX96 real-time PCR system C1000 thermal cycler(Bio-Rad, Hercules, CA, USA) 기기를 사용법에 맞게 실험을 진행하였다. PCR에 사용된 primer는 MUC5AC primer(QT01329615, Qiagen, Hilden, Germary) and MUC5B primer(BioRad 10025636, qHsaCIP0028135, Hercules, CA, USA)에서 구입하여 사용하였으며 각 반응의 내부 양성 대조군(internal positive control)은 glyceraldehyde-3-phosphate dehydrogenase(GAPDH)를 사용하였다. GAPDH의 경우 forward는 5’-CCT CCA AGG AGT AAG ACC CC-3’, reverse는 5’-AGG GGT CTA CAT GGC AAC TG-3’을 사용하였다.
이 과정을 간략히 설명하면, 배양된 세포를 2% bovine serum albumin을 함유한 PBS로 3회 세척한 후 TrizolⓇ(Molecular Research Center, Cincinnati, OH, USA)을 이용하여 총 mRNA를 추출하였다. Reverse transcription PCR을 진행하여 cDNA를 합성한 후, Real-time PCR을 진행하였다. iQ SYBR Green Supermix 사용하여 95°C에서 15초간 변성과정을 거치고 60°C에서 45초간 결합반응을 시키는 과정을 50회 반복하였다. 증폭의 정확도는 변성곡성(melting curve, Roche Applied Science, Mannheim, Germany)을 사용하여 평가하였다.
면역분석법(Immunoassay)
MUC5AC과 MUC5B의 점액 단백의 함량을 측정하기 위해서 enzyme-linked immunosorbent assay(ELISA)법을 이용하였다. 시료를 처리한 배양된 세포에서 radioimmunoprecipitation assay buffer(Thermo Scientific, Rockford, IL, USA)로 단백을 추출하여 BCA를 이용하여 정량 분석 하였다. 추출한 단백질 20 μg을 F96 Cert. Maxisorp Nunc-Immuno plate(Fisher scientific, Lenexa, KS, USA)에 4°C에서 24시간 배양시킨 후, plate를 PBS로 3회 세척하였다. 비특이적 결합을 방지하기 위해 2% bovine serum albumin으로 실온에서 1시간 동안 배양한 후, PBS로 3회 세척한 다음 0.05% Tween 20을 함유한 PBS에 1:200으로 희석된 MUC5AC, MUC5B 일차항체로 반응시켰다. 다시 PBS로 3회 세척한 후 HRP-conjugated 이차항체를 0.05% Tween 20을 함유한 PBS에 1:5,000으로 희석하여 각 well에 첨가하였고, 1시간 후에 각 well을 PBS로 3회 세척하였다. 3,3’, 5,5’-tetramethyl benzidine 용액으로 발색한 후, 2N-H2SO4를 이용하여 중단시켰다. EL800 ELISA reader(BIO-TEK Instruments, Winooski, VT, USA)로 450 nm에서 흡광도 측정 후 표준곡선을 이용하여 단백의 양을 정량화하였다.
결 과
NCI-H292 세포에서 담배특이니트로사민의 세포 생존에 미치는 영향
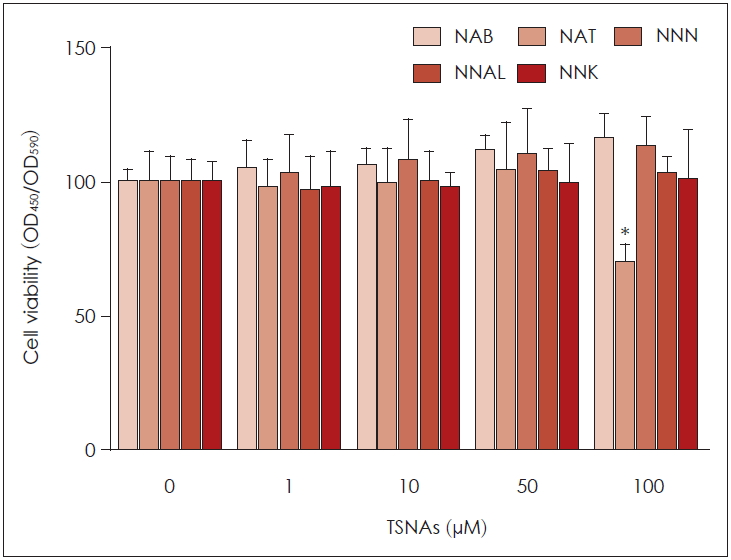
NCI-H292 세포에 담배특이니트로사민인 NAB, NAT, NNN, NNAL과 NNK를 처리하고 WST-1을 이용하여 세포 생존에 미치는 영향을 확인한 결과, NAB, NNN, NNAL과 NNK는 100 μM까지 세포생존에 영향을 주지 않았다. 하지만 NAT는 100 μM에서 세포생존율이 70%로 통계적으로 유의미하게 감소하였다(Fig. 1).
NCI-H292 세포에서 담배특이니트로사민이 MUC5AC mRNA와 단백 생성에 미치는 영향
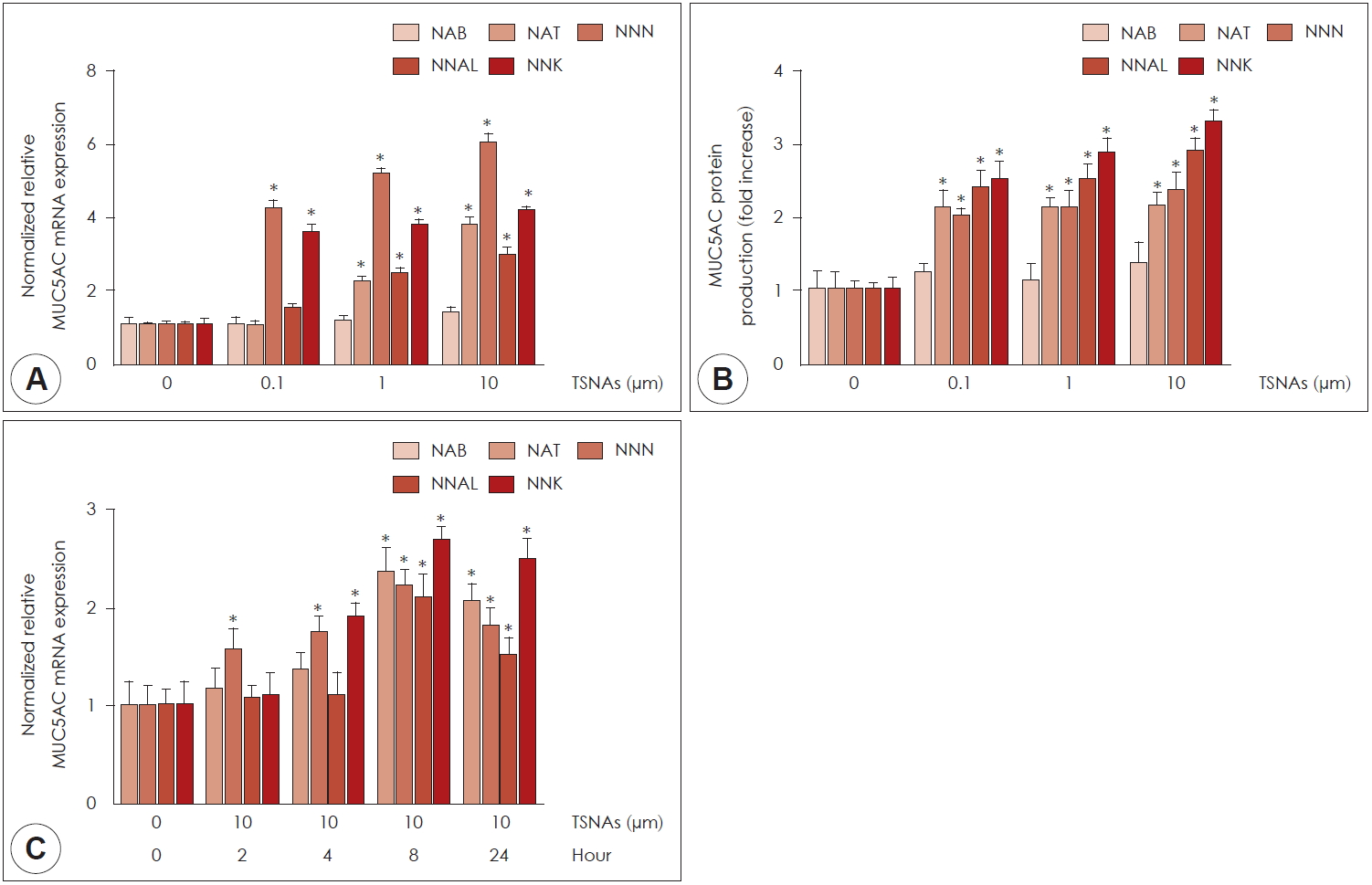
담배특이니트로사민인 NAB, NAT, NNN, NNAL과 NNK를 0.1, 1, 10 μM 처리한 후 real-time PCR및 ELISA를 시행한 결과, NAT, NNN, NNAL과 NNK에 의해 점액유전자 MUC5AC mRNA 발현과 단백 생성이 의미있게 증가하였다(Fig. 2A, B). NAB는 점액유전자 MUC5AC mRNA 발현과 단백 생성 모두 농도에 따른 통계학적인 차이를 나타내지 않았다. 따라서 NAB는 추후 실험에서 제외하였다. 시간에 따른 NAT, NNN, NNAL과 NNK에 대한 MUC5AC mRNA 발현 정도를 알아보기 위해 NAT, NNN, NNAL과 NNK을 10 μM 농도로 2, 4, 8, 24시간 동안 처리하여 real-time PCR을 시행하였다. NAT, NNN, NNAL과 NNK의 투여에 대한 MUC5AC mRNA 발현이 증가하였고 NAT, NNN, NNAL, NNK 투여 8시간 후에 최고치에 도달하였다(Fig. 2C).
NCI-H292 세포에서 담배특이니트로사민이 MUC5B mRNA와 단백 생성에 미치는 영향
담배특이니트로사민인 NAB, NAT, NNN, NNAL, NNK를 0.1, 1, 10 μM을 처리한 후 real-time PCR 및 ELISA를 시행한 결과, NAB, NAT, NNN, NNAL, NNK에 의한 MUC5B mRNA 발현과 단백 생성 모두 유의미한 차이를 나타내지 않았다(Fig. 3).
NCI-H292 세포에서 NAT, NNN, NNAL, NNK이 MUC5AC mRNA와 단백 생성에 관여하는 수용체 확인
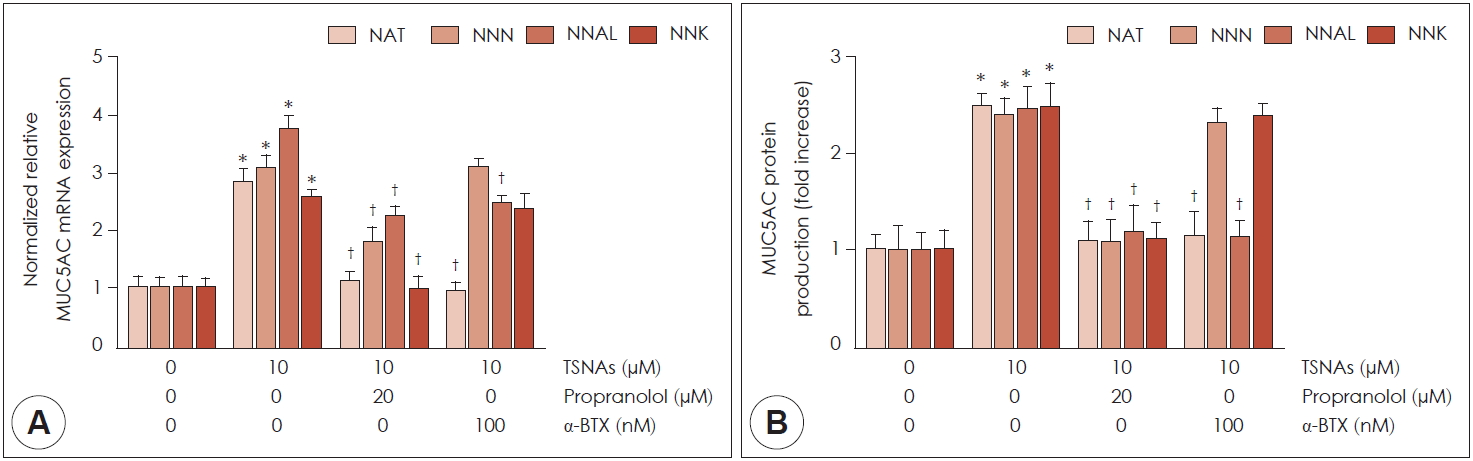
NCI-H292 세포에서 MUC5AC의 mRNA 발현과 점액단백의 생성에 관여하는 수용체의 여부를 확인하기 위해 담배특이니트로사민과 관련있는 2가지 수용체의 억제제를 이용하였다. Propranolol(β-adrenergic receptor antagonist, 20μM)와 α-bungarotoxin(α-BTX, a α7-nicotinic acetylcholine receptor antagonist, 100 nM)를 2시간동안 세포에 전처리 한 뒤, NAT, NNN, NNAL과 NNK를 처리하여 real-time PCR 및 ELISA를 시행하였다. NAT, NNAL에 의해 증가된 MUC5AC의 mRNA 발현과 단백 생성은 Propranolol과 α-BTX에 의해 모두 통계학적으로 유의미하게 억제되었다(Fig. 4). 하지만 NNN과 NNK에 의해 증가한 점액유전자 MUC5AC mRNA 발현과 단백생성이 propranolol에 의해서는 통계학적으로 유의미하게 억제 되었지만 α-BTX를 처리한 군에서는 효과적으로 억제 되지 않았다(Fig. 4).
고 찰
전자담배(E-cigarette)는 주로 니코틴이 들어있는 액상을 기기에서 기화시켜 수증기를 흡입하는 형태로 일반담배(tobacco cigarette)에 비해 해롭지 않다고 알려져 있다[18]. 하지만 니코틴이 고온·고압에서 기화되어 니코틴 부산물인 니트로사민이 발생하면 각종 질환을 일으킬 수 있다[19]. 니트로사민의 배출 함량은 제품에 따라 다르지만, 일반담배의 경우 평균 96 ng/1puff이 검출되었고, NNN은 53 ng, NAT는 41ng였고 NAB, NNK는 검출되지 않은 제품이 대다수였다. 전자담배의 경우는 니트로사민이 평균 0.22 ng/1puff이 검출되었고 NNN은 0.01 ng, NAT는 0.1 ng, NAB는 0.03 ng, NNK는 0.09 ng이 검출되었다. 또한 니코틴액 21가지 제품에서 TSNAs의 농도를 측정한 결과 농도 범위가 37.8~2324.5 ng/mL이며, 전자담배의 기기의 종류나 니코틴액의 제품마다의 차이가 나고 전압과 저항도에 따라 니트로사민의 배출량의 차이가 생기기 때문에 전자담배에서 생성된 담배특이니트로사민의 위험성을 역시 간과할 수 없다[20]. 담배특이니트로사민은 폐로 들어가 정상세포에 작용하여 정상세포를 암세포로 전환시키면서 폐암 발생에 관여하고 만성폐쇄성폐질환과 폐섬유화 등과 같은 폐질환이 있는 경우에는 폐암 발병 위험을 높이는 것으로 알려져 있다[21]. 하지만 호흡기에서 점액유전자 발현에 미치는 영향에 대해서는 알려진 바가 거의없다.
대표적인 염증성 호흡기 질환인 천식(asthma)이나 만성폐쇄성폐질환(Chronic obstructive pulmonary disease, COPD), 만성 기관지염에서 점액의 과분비는 중요한 병인 중 하나이다[22]. 염증반응이나 감염에 의해 점액이 과분비 되면 점액섬모운동에 이상이 생기고, 그로 인해 점액섬모 청소기능이 떨어져 질병을 악화시켜 병의 경과에 악영향을 미친다[8]. 따라서 본 연구는 호흡기 상피세포에서 니코틴 부산물인 담배특이니트로사민(NAB, NAT, NNN, NNAL, NNK)에 의한 점액유전자 발현과 단백 생성을 알아보고 관련 수용체에 대해 알아보고자 하였다.
연구결과 NAT, NNN, NNAL, NNK에 의해서 MUC5AC유전자 발현과 단백 생성이 통계학적으로 유의미하게 증가하였다. 하지만 NAT, NNN, NNAL과 NNK는 MUC5B발현에 영향을 주지 않았다. 또한, NAB는 MUC5AC와 MUC5B의 발현에 모두 통계학적 유의미한 증가를 나타내지 않았다. MUC5AC 와 MUC5B는 젤형성에 관여하는 점액유전자로 대표적인 분비형 점액유전자이다. 천식환자 군에서 MUC5AC와 MUC5B의 유전자 발현 양상이 변하고 천식 발병기전에 영향을 미친다[23]. 낭포성 섬유증(cystic fibrosis) 환자군에서는 MUC5AC와 MUC5B가 과발현 되어 있다[24]. 하지만 MUC5AC와 MUC5B는 형태학적 및 생물리학적 특성이 다르기 때문에 다른 경향이 나타날 수 있다. MUC5AC는 배상세포(goblet cell)의 과형성 표식인자로 주로 배상세포에서 분비되고, MUC5B는 주로 점막하선에서 분비된다[9]. MUC5AC는 여러 염증반응과 감염질환에서 증가되어 있는 경향이 있다. MUC5AC와 달리 MUC5B는 상기도의 감염 조절에 영향을 미치고 하기도의 면역 항상성 조절에 관여한다[25]. 또한 MUC5AC가 결핍된 실험동물과 달리 MUC5B의 결핍된 실험동물에서는 점액섬모운동에 이상이 나타나고 박테리아 감염률이 증가하는 현상으로 인해 치사율이 높게 나타난다[25]. COPD환자군의 경우, MUC5AC의 발현은 질환 중증도에 따라 불균형하게 증가하지만, MUC5B는 질병의 중증도에 따라 증가하였다[26]. 점액유전자의 생물학적 역할이 다른 이유로 인해 담배특이니트로사민의 구조나 특징에 따라 발현의 차이가 나타난다고 예상된다.
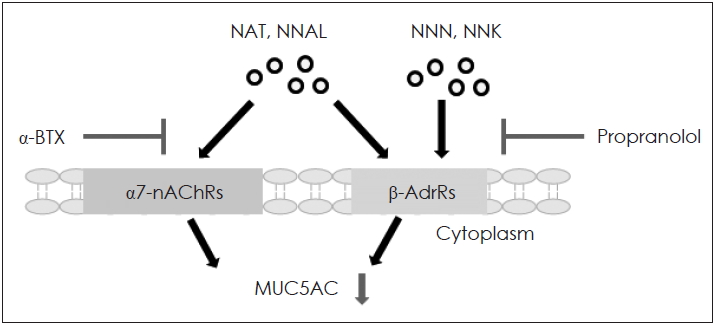
니코틴 부산물인 니트로사민은 세포 표면에 있는 α7-nicotinic acetylcholine receptor(α7-nAChRs)과 β-adrenergic receptor(β-AdrRs)을 통해 생물학적 효과를 나타낸다[27]. α7-nAChRs는 주로 상피세포에 존재하며 폐에서 니코틴 또는 니트로사민과 결합하여 다양한 염증반응을 매개한다고 알려져 있다[28]. 니코틴은 α7-nAChRs을 통해 mucous cells의 metaplasia/hyperplasia를 유도하고 점액유전자 MUC5AC와 MUC5B를 증가시킨다[29]. 또한 약물작용과 중추신경계에 관여하고 이온 체널 영향을 주어 양이온(Na+, Ca2+, K+)의 흐름을 조절 하는 등 다양한 생물학적 약리학 및 병리 생리학적 역할을 수행한다[27]. 따라서 이러한 수용체의 조절과 관찰은 질병을 예방하고 치료하는데 중요한 역할을 할 수 있다. β-AdrRs은 G protein-coupled receptor로 인체에서 세포 외부의 자극을 세포 내 신호전달하는 역할을 한다[30]. β-AdrRs는 사람 호흡기 NCI-H292 상피세포에서 cigarette smoke extract에 의해서 점액을 과발현 시킨다고 알려져 있으며, 억제제인 propranolol에 의해 억제된다[31]. 따라서 본 연구진은 니트로사민의 수용체인 α7-nAChRs과 β-AdrRs의 억제제를 이용하여 점액유전자와의 관계를 확인해 보았다. α7-nAChRs 억제제인 α-BTX과 β-AdrRs의 억제제인 propranolol를 사용한 결과, propranolol과 α-BTX에의해 NAT, NNAL에 의해서 증가한 점액유전자 MUC5AC mRNA와 단백생성을 모두 효과적으로 억제시켰다. 하지만 NNN과 NNK에 의해서 증가한 MUC5AC는 propranolol에 의해서만 유의미하게 억제 시켰으며 α-BTX에 의해서는 통계학적으로 의미 있는 억제를 시키지 못했다. NNN과 NNK가 β-AdrRs에 작용하여 MUC5AC 과발현에 영향을 주지만 α7-nAChRs를 통해서는 MUC5AC 발현에 관여하지 않을 것으로 예상된다(Fig. 5). 이 결과들은 담배특이니트로사민이 호흡기 상피세포에서 α7-nAChRs그리고/또는 β-AdrRs를 통해 점액유전자 MUC5AC의 발현을 증가시킨다는 것을 제시할 수 있다. 하지만 담배특이니트로사민이 점액유전자의 과분비 효과를 명확히 확인하기 위해서는 수용체의 하위 기전을 밝히고 다른 종류의 세포실험 및 동물실험을 통해 검증하는 연구가 뒤따라야 할 것으로 생각된다.
결론적으로 본 연구는 사람 호흡기 상피세포에서 담배특이니트로사민인 NAT, NNN, NNAL과 NNK로 인해 점액유전자 MUC5AC 발현을 증가 시켰다. 또한 TSNAs의 특이적 수용체인 α7-nAChRs과 β-AdrRs에 의해 점액유전자가 증가하였으며 담배특이니트로사민의 구조적 차이에 따라 점액유전자 발현양상이 다르게 나타났다. 따라서 담배 또는 전자담배로 흡연으로 인한 TSNAs의 노출이 증가하면 점액유전자가 과발현되어 호흡기 질환의 악화에 영향을 줄 것으로 생각되며 이와 관련된 추가적인 연구가 필요한 것으로 생각한다.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print