INTRODUCTION
The incidence of allergy and asthma has rapidly increased during the past few decades, and these disorders are now some of the most common chronic diseases worldwide [1,2]. Asthma is a chronic inflammatory disorder caused by an imbalance between the Th1 and Th2 [3,4]. Asthmatic symptoms arise from the cross-linking of allergen with antigenspecific IgE molecules that bind to mast cells, causing their activation and degranulation, and release of histamine, leukotrienes, and other inflammatory substances. This effect causes recruitment of eosinophils, which are regulated by interleukin-5, and cause airway damage and hyper-responsiveness [5,6].
The role of the gut microbiota in gastrointestinal (GI) development, immune adaptation, and attenuation of inflammatory diseases of the GI tract is of critical concern. Moreover, there are steadily convincing evidences that certain commensal bacteria could regulate immune responses outside the GI tract. Notably, there is a rising interest in the therapeutic effects of the microbiome in allergic disorders [7,8].
Bacillus clausii (BC) is a probiotic widely used in Italy since the 1960s for viral diarrhea in children and antibiotics-associated diarrhea. Clinical trials have shown that BC is useful in the management and prevention of recurrent respiratory infections in children [9]. This probiotic has been demonstrated to exert immunomodulatory activities. However, few studies have examined the anti-allergic effects of BC in individuals with allergic diseases.
Recent studies have shown that oxidative stress can mediate a significant effect on the pathogenesis of allergic diseases [10]. Antioxidants can attenuate asthma by suppressing IgE, Th2 cytokines, mucus hypersecretion, and eosinophilic infiltration in a murine model. Moreover, antioxidant agents decrease ROS generation and blockade of airway hyperresponsiveness and inflammation via regulation of NF-╬║B and HIF-1╬▒ levels [11].
Some probiotics have been identified as antioxidative, as they contain high levels of glutathione and Mn-SOD. Probiotics can improve the Th1/Th2 balance, and intracellular levels of some antioxidant enzymes, including glutathione peroxidase and SOD [12]. However, it is unclear whether the antioxidative signaling of probiotics including BC, could play a role in attenuating allergic airway inflammation.
In this study, we demonstrated that administration of BC attenuated allergic airway inflammation through regulation of the hypoxia signaling pathway, whose effect is quite comparable to that of Lactobacillus paracasei (LP) administration, one of the most well-known probiotic strains.
MATERIALS AND METHODS
Animals
Six-week-old female BALB/c mice (18-20 g) were used (Orient Bio, Seongnam, Korea). All mice were raised in a specific-pathogen-free environment with a 12-hour daynight cycle, and received allergen-free water and food. They were also able to move freely without restriction. All experiments were performed according to the protocol approved by the Institutional Animal Care and Use Committee of the Institute (IACUC, approval number INHA 180303-541).
Induction of allergic asthma/administration of LP or BC probiotics
The mice in the control group were sensitized and challenged with normal saline. For the induction of allergic asthma, mice in the OVA group were sensitized with an intraperitoneal (i.p.) injection of 20 ╬╝g OVA (Sigma-Aldrich, St. Louis, MO, USA) and 2.25 mg aluminum hydroxide gel (as an alum adjuvant: Thermo, Waltham, MA, USA) in 100 ╬╝L saline on days 0 and 14. After systemic sensitization, mice were challenged by intranasal instillation of OVA (500 ╬╝g) on days 28, 29, and 30 (Fig. 1).
LP was obtained from the Korean Collection of Type Cultures, Republic of Korea. BC was isolated from the tidal mudflats of the Korean Yellow Sea and was screened using a skimmed milk agar plate in tryptic soy broth. Based on the 16S rRNA sequence data, we identified the isolate I-52 as B. clausii, as it had high homology with B. clausii DSM 8716T. We have deposited LP in the Korean Collection for Type Cultures (Daejeon, Republic of Korea), under the accession #KCTC 10277 BP.
We diluted LP or BC in normal saline and administered it orally, every day from the week before the first sensitization to the last OVA challenge. Mice in the LP1 or BC1 group were administered 0.2 mL of LP or BC at doses of 1├Ś109. CFU/mL. Mice in the LP5 or BC5 group were administered 0.2 mL of LP or BC, at doses of 5├Ś109 CFU/mL, respectively (Fig. 1).
Measurement of differential cell counts and titers of Th2 cytokines in bronchoalveolar lavage fluid (BALF)
One day after the last intranasal challenge with OVA, mice were sacrificed, and the trachea was immediately cannulated with an intravenous polyethylene catheter equipped with a 20-gauge needle on a 1-mL syringe. Lungs were washed twice with 0.8 mL PBS. The BALF was spun down, the supernatant was removed for ELISA, and the pelleted cells were resuspended in PBS for counting the BALF cells. Cytospin was prepared from resuspended BALF cells by centrifuging the cells onto microscope slides. Slides were stained with Diff-Quick staining (Thermo Electron Corporation, Pittsburg, PA) and quantified for differential cell counts by counting 100 cells/slide at ├Ś40 magnification. We measured the titer of cytokines IL-4 and IL-5 using ELISA kits (BD Bioscience San Diego, CA, USA), according to the manufacturerŌĆÖs instruction.
Histopathology of lung tissue
The whole left lung tissues of mice were fixed in 10% neutral buffered formalin, embedded in paraffin, cut into 3-mm sections, and were stained with hematoxylin and eosin (H&E) and periodic acid-Schiff (PAS) (Diagnostic Biosystems, CA, USA) to evaluate the degree of goblet cell hyperplasia in the bronchial epithelium. We used a light microscope (Olympus Bx43, Olympus Corporation, Tokyo, Japan) for pathological analysis.
Measurement of the serum total and OVA-specific IgE
We collected blood from the abdominal aorta by an aortic puncture technique. Whole blood was centrifuged for 30 minutes, at 4┬░C, 13,000├Śg, and the supernatant (serum) was stored at -80┬░C until further analysis. We measured the titer of serum total IgE using an ELISA kit (BD Bioscience San Diego, CA, USA), according to the manufacturerŌĆÖs instructions.
To evaluate OVA-specific IgE and IgG1a in the serum, OVA (10 ╬╝g/well) was coated in the 96-well plates overnight at 4┬░C. The next day, the solution was removed, the wells were washed four times, and 200 mL of blocking solution was added and incubated at 37┬░C for 1 h. The dilutions of serum with sample diluent were then added to the wells and incubated at 37┬░C for 2 h. The plates were washed four times with PBST, and biotin-anti IgE (BD Bioscience) or biotin-anti IgG1 (BD Bioscience) antibody was added to the wells and incubated at 37┬░C for 1 h. After washing, we added 100 mL of diluted horseradish peroxidase (HRP)- conjugated antibody to each well, and the plates were incubated again at 37┬░C for 30 min. The plates were washed again four times, and we added 100 mL of tetramethylbenzidine (TMB) substrate solution to each well. After that, we kept the plates in the dark at room temperature for color development. The enzyme reaction was stopped by adding 100 mL of stop solution. Absorbance was measured at 450 nm using an ELISA plate reader.
Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) analysis
Whole-lung tissue was frozen in liquid nitrogen immediately after harvesting and was homogenized in 1 mL TRIzol reagent (Invitrogen Life Technologies, Waltham, MA, USA). Samples were stored at -80┬░C until further analysis. We isolated total RNA as recommended by the manufacturer. In total, 1 ╬╝g of RNA was subjected to reverse transcription using a PrimeScript First Strand cDNA Synthesis Kit (Takara Bio Inc., Shiga, Japan). We then performed Real-time PCR in duplicate using the SYBR Green Master Mix ABI Prism on a PCR machine (StepOne plus Real-Time PCR System; Applied Biosystems Inc., Carlsbad, CA, USA). The mRNA expression of HO1 and Hif1╬▒ was detected using Mm_Hmox1_1_SQ and Mm_ Hif1╬▒ _1_SQ QuantiTect Primer Assays (Qiagen, Valencia, CA, USA). The housekeeping gene glyceraldehyde-3-phosphate dehydrogenase was used as an internal control to normalize gene expression.
Immunohistochemistry
Lung paraffin sections were deparaffinized and hydrated using xylene and a graded alcohol series. Antigen retrieval was performed with citrate, buffer pH 6.0, at 95┬░C for 20 min (DAKO, CA, USA). Endogenous peroxidase activity was blocked with 2.5% hydroxyl peroxide-diluted 1.5% normal blocking serum for 20 min at room temperature. For immunohistochemistry, the Vecta Stain Elite ABC kit (Rabbit IgG, Vectorlabs, PK-6101) was used according to the manufacturerŌĆÖs protocol, and the following primary antibodies were used: HO1 (Abcam, ab13243, 1:500), Hif1╬▒ (Abcam, ab82832, 1:1000). On the next day, sections were washed three times with PBS for 5 min each, and then incubated with a secondary antibody: biotinylated goat antirabbit IgG (Vector Laboratories) for 1 h at room temperature. Vectastain Elite ABC reagent (Vector Laboratories) was applied to the sections for 30 min at room temperature After washing with PBS, the ImmPACT DAB peroxidase (HRP) substrate kit (Vector Laboratories) was used according to the manufacturerŌĆÖs protocol for color development. After rinsing with tap water, nuclei were counterstained with hematoxylin (Sigma-Aldrich) and the sections were then dehydrated through a graded alcohol series and xylene.
Statistical analysis
WE expressed results as mean┬▒standard deviation (SD). Statistical differences were assessed using the KruskalWallis test or Mann-Whitney U test. For multiple comparisons, the Kruskal-Wallis test was used. As significant differences were found, Mann-Whitney test was used to compare statistical differences between two groups. Statistical analyses were performed using GraphPad Prism 4.01 (GraphPad Software, La Jolla, CA, USA). P-values<0.05 were considered statistically significant.
RESULTS
Administration of probiotics reduced allergic pulmonary inflammation
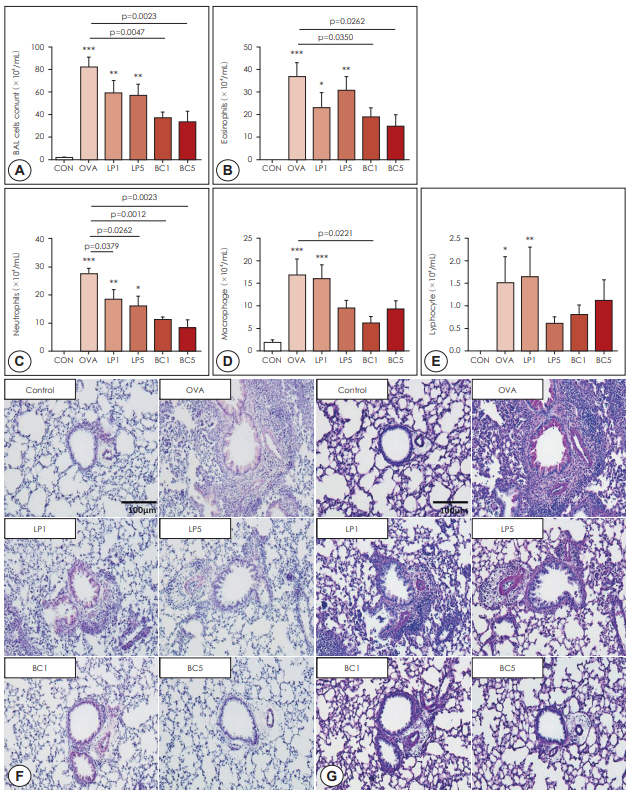
By analyzing differential cell counts in BALF, we observed that the numbers of total inflammatory cells (p<0.001), eosinophils (p<0.001), neutrophils (p<0.001), macrophages (p<0.001), and lymphocytes (p<0.05) were all significantly increased in the OVA group compared to the control group. After oral administration of LP or BC, the number of eosinophils, neutrophils, and lymphocytes was considerably reduced compared to that in the OVA group (p<0.05, Fig. 2A-E). Compared to those in the LP group, mice in the BC group exhibited substantially lesser BALF eosinophils (LP5 versus BC5: p=0.008) and neutrophils (LP5 versus BC5: p=0.047).
Histopathologic examination using H&E staining showed that infiltration of inflammatory cells and thickening of the airway epithelium was significantly reduced after LP or BC treatment (Fig. 2F). Secretion of mucus in the airway epithelium was observed by PAS staining. After LP or BC treatment, the number of PAS-positive cells was significantly decreased compared to that in the OVA group (Fig. 2G). These results suggest that probiotics attenuated inflammatory cell infiltration and mucus secretion.
Administration of probiotics affected Th2 immune response
We measured the effect of probiotics on total IgE, a typical indicator of asthma in the serum. In the LP or BC-treated group, total IgE levels tended to be decreased compared to that in the OVA group, although without statistical significance (p>0.05, Fig. 3A). OVA-specific sIgE levels did not show a significant difference between groups (p>0.05, Fig. 3B).
On the contrary, OVA-specific IgG1 level was significantly decreased in the LP and BC groups, compared to the OVA group (p<0.05, Fig. 3C).
IL-4 (p<0.001) and IL-5 (p<0.001) were significantly increased in the OVA group, compared to the control group. After LP or BC treatment, the titer of IL-4 and IL-5 was significantly decreased compared to that in the OVA group (p<0.05, Fig. 3D, E). Especially, BC1 and BC5 groups showed a significant decrease in the IL-4 titer compared to the LP group (p=0.045).
Administration of probiotics suppressed HIF-1╬▒ signaling
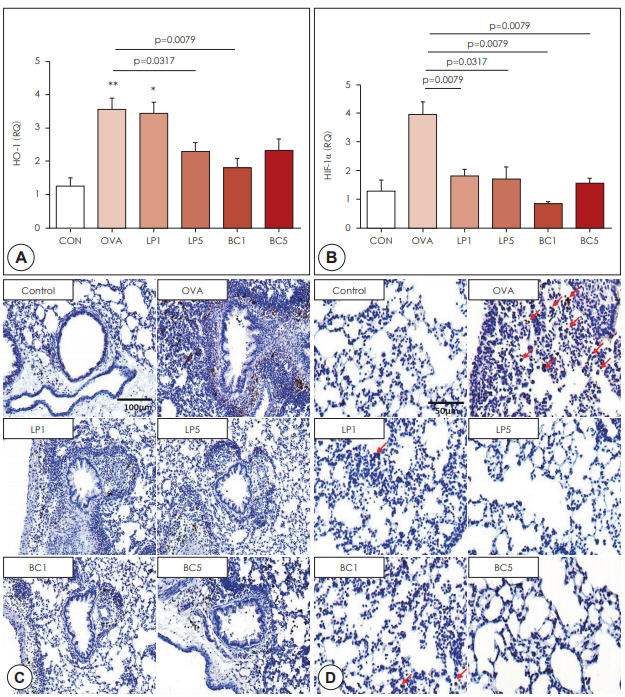
To determine whether probiotics affect the hypoxia-related pathway in asthmatic lungs, we examined the expression of HO-1 and HIF-1╬▒ by real-time PCR. HO-1 and HIF-1╬▒ expression was significantly upregulated in the OVA group, compared to the control group (p<0.001, Fig. 4A, B). After oral LP or BC treatment, the mRNA expression of these genes was significantly suppressed compared to that in the OVA group.
Increased HO-1 (p<0.001) and HIF-1╬▒ (p<0.001) mRNA expression in the OVA group was significantly decreased in the probiotic groups (p<0.05, Fig. 4A, B). Using immunohistochemical staining, we found that HO-1 and HIF-1╬▒ protein expression in the LP or BC group was reduced significantly, compared to that in the OVA group (Fig. 4C, D).
DISCUSSION
The novel findings of the our study were as follows: 1) administration of LP or BC attenuated airway inflammation (manifested by decreased inflammatory cells in BALF) and asthma (proven by improved histopathologic findings); 2) the probiotics reduced Th2 immune responses by reducing the production of IL-4, IL-5, and OVA-specific IgG1; and 3) the blockage of HO-1- HIF-1╬▒ axis by probiotics was at least partially responsible for the asthma improvement.
The total cell count in BAL fluids and the percentage of eosinophils was significantly lower in LP or BC treatment groups compared to that in the allergy group. Histopathological analysis revealed that peribronchial infiltration of inflammatory cells was significantly decreased by treatment with LP or BC. These results are in agreement with those from previous studies, demonstrating that probiotics reduce inflammatory cell numbers in BAL fluids, and inflammatory cell infiltration into lung tissues in animal models of asthma [13,14]. These results indicate that the new mudflat-derived probiotic BC, has anti-allergic effect comparable to that of the well-known probiotic LP.
In this study, serum IgG1 levels were significantly lower in the LP or BC treatment groups compared with the asthma group. Considering these results, we believe that the Th2 immune response is modified by treatment with LP or BC. This is consistent with the results reported by Robinson et al. [15] that elevated allergen-induced Th2 type cytokine production in the airway leads to increased Th2 immune responses and eosinophilic airway inflammation. The mechanism by which probiotics such as LP or BC regulate allergic diseases may be through restoration of Th1/Th2 imbalance.
In the present study, our results showed that OVA-sensitized asthmatic mice expressed higher levels of IL-4 and IL-5 in the lung than control mice, but the IL-4 and IL-5 levels decreased after treatment with LP or BC. Lowering of IL-4 and IL-5 production is related to the features of asthma, responsible for the decrease in BALF eosinophil infiltration, IgE production, and Th2 immune response [4].
Probiotics promote potentially anti-allergic and anti-inflammatory processes [16]. Recent studies have shown that BC has stronger suppressive effects on the development of allergic inflammation [17]. As for in vivo experiments, previous studies have already demonstrated that LP has an immunosuppressive effect on OVA-sensitized asthmatic mice [18]. Therefore, this study aimed to explain the effectiveness of BC and its possible mechanism in the immunosuppressive response in vivo.
BC is considered safe for all ages and is effective in the treatment of several diseases. In particular, Bacillus clausii has also been shown to exert various health effects on the immune system. Its immune regulatory activity relates to an increased percentage of T lymphocytes as a marker of T-cell activation.
There are several similarities between studies conducted with other probiotics and the current study. Both studies have shown reduced AHR and lung inflammatory status. However, studies with other probiotics did not demonstrate significant decreases in the number of eosinophils in BALF and focus on Th2 immune response, as shown in this study. Different probiotic treatments or allergic diseases might produce different results, but the mechanisms of probiotic therapy are becoming clear.
Furthermore, our study indicated that a reduction in AHR is related to the decrease of serum immunoglobulin production and less eosinophil infiltration in OVA-sensitized mice after oral probiotic administration. Our results also suggest that the anti-allergic effect in the BC group was quite comparable to that in the LP group. Further, decreased IL-4 and IL-5 production in lung tissues was associated with reduced eosinophil infiltration in Bacillus clausii-treated mice, suggesting that Bacillus clausii plays a significant role in suppressing the asthmatic responses in OVAsensitized mice. In short, this study suggests a possible mechanism for the immunosuppressive effects of Bacillus clausii in modulating the Th2 pathway mediated by HIF-1╬▒ signaling in an animal model.
In this study, we identified a novel regulatory mechanism in OVA-induced airway inflammation wherein downregulating probiotics and further suppressing the expression of HIF-1╬▒ results in a decrease in HO-1, the anti-inflammatory protein. HIF-1╬▒ is a transcription factor, playing critical roles in modulating innate immunity in the lung and regulates downstream genes such as HO-1 by binding to hypoxia-responsive factors [19,20]. When the lungs are stimulated with OVA, reactive oxygen species are generated for eradicating other pathogens [21]. It was demonstrated that ROS upregulated the induction of HIF-1╬▒ expression involved in hypoxia signaling.
In summary, this study demonstrates that Bacillus clausii alleviates asthmatic symptoms in a murine model of asthma. Furthermore, we found that Bacillus clausii might play a role in the HIF-1╬▒ pathway-mediated pulmonary allergic inflammation, which probably results in immune-regulatory properties. An in-depth study of this suppressive effect should lead to an increased understanding of the features of Bacillus clausii and an expansion of its clinical therapy.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print